
J. M. Pearce1, J. Dooley2, V. Patil1, T. L. Sformo3,4, B. L. Daniels5, A. Greene6, and J. Leafloor7
1Alaska Science Center, U.S. Geological Survey, Anchorage, AK, USA
2Division of Migratory Bird Management, U.S. Fish & Wildlife Service, Vancouver, WA, USA
3Department of Wildlife Management, North Slope Borough, Utqiaġvik (Barrow), AK, USA
4Institute of Arctic Biology, University of Alaska Fairbanks, Fairbanks, AK, USA
5Yukon Delta National Wildlife Refuge, U.S. Fish & Wildlife Service, Bethel, AK, USA
6Kotzebue, AK, USA
7Aquatic Unit, Environment and Climate Change Canada, Winnipeg, MB, Canada
Highlights
- Multiple species of geese spend part of their annual cycle in the circumpolar Arctic and serve as a source of nutrition and cultural affirmation for many peoples.
- Arctic geese function as important indicators of environmental changes and some species also have the potential to alter ecosystem processes when they become overabundant.
- In 2022, despite an outbreak of highly pathogenic avian influenza in North America and variable spring weather conditions, the population sizes of most Arctic geese remained at or above historical levels.
Introduction
Most species of geese in the Northern Hemisphere are long-distance migrants, moving between lower-latitude wintering areas and summer breeding areas in the Arctic and sub-Arctic (Fig. 1). Arctic geese (Fig. 2) are keystone herbivores and their numbers and distributions are influenced by summer and winter environmental conditions and changes in forage quality and quantity. Geese are one of the first avian species to return to northern regions in early spring and these birds and their eggs are highly valued as a source of nutrition and cultural affirmation for Indigenous Peoples and Arctic residents.
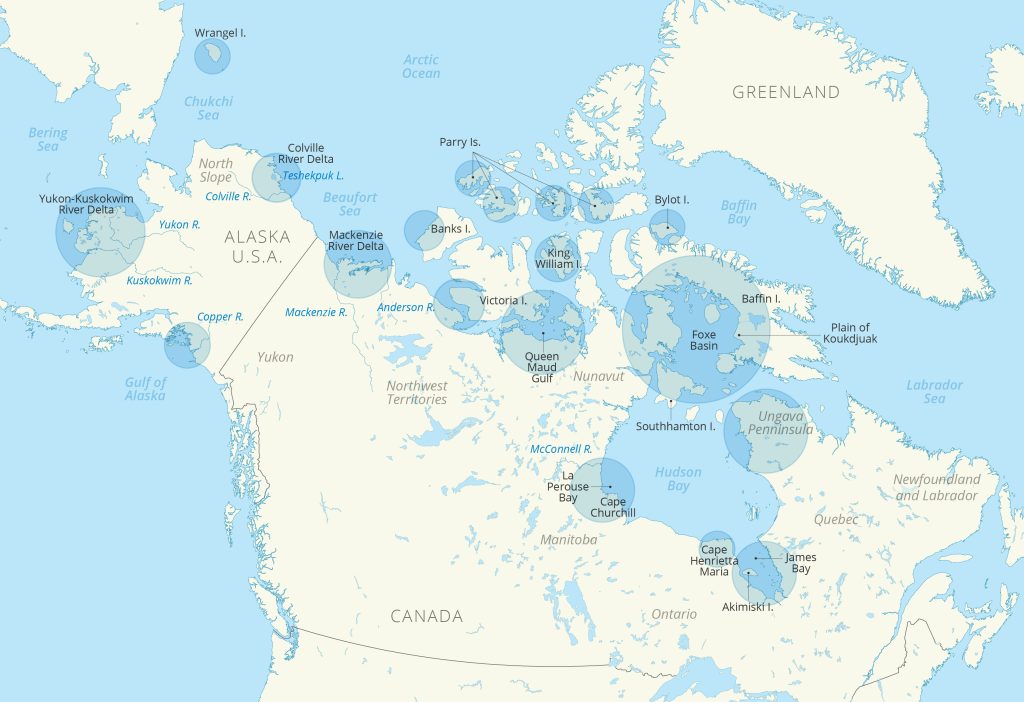

This essay focuses on the status of North American goose populations that breed within areas of the Arctic and sub-Arctic (Fig. 1). Within North America, the status and trends of waterfowl species and their breeding habitats are of significant interest to federal, state, and tribal entities and are primarily assessed through standardized ground and aerial surveys and the marking of individuals (U.S. Fish and Wildlife Service 2022). Additionally, the Arctic Goose Joint Venture (AGJV) publishes a strategic plan that summarizes information needs across North America (https://www.agjv.ca) and provides links to current species and population management plans. These efforts, along with annual coordination among agencies and co-management councils, documentation of Inuit Knowledge of Arctic geese (e.g., www.kangut.ca), and involvement of Indigenous Peoples and Arctic residents, help to provide the most up-to-date status information, ensure sustainable harvest regulations, and improve our collective knowledge of these birds. An in-depth review of the abundance, status, and distribution of circumpolar Arctic geese can be found in Fox and Leafloor (2018).
Spring conditions in 2022
An outbreak of highly pathogenic avian influenza (HPAI) that began in both eastern Canada and British Columbia and quickly spread across North America (U.S. Geological Survey 2022) affected many Arctic geese in 2022. This outbreak, only the second ever detected in North America, has already resulted in more losses of wild and domestic birds than the previous outbreak during 2014-16. HPAI mortality events in 2022 occurred across all migratory flyways during the spring migration period and involved notable die-offs of Canada geese and lesser snow geese in some staging areas where they rest and feed along their migration. HPAI was also detected in several species of migratory geese in breeding areas of Alaska from early May 2022 to August 2022 (Alaska Division of Environmental Health 2022). Live geese were swabbed during banding in the Canadian Arctic in 2022 as part of national HPAI surveillance activities, and no active mortality events were observed at that time; further analysis of these samples is currently underway.
In Canada, biologists noted favorable spring 2022 conditions that led to average to above-average goose productivity (i.e., young geese observed) on Banks Island, Southampton Island, Baffin Island, the Ungava Peninsula, and along western Hudson Bay (Fig. 1). However, low production was noted for geese in some areas of the central Arctic and on Bylot Island. Spring phenology was later than average on Bylot Island but appeared to be early or average in many areas of the Canadian Arctic and sub-Arctic. Many areas in these regions also experienced above-average temperatures during late May or June (U.S. Fish and Wildlife Service 2022; see similar observations in essay Surface Air Temperature, Figs. 2c and 2d, and essay Terrestrial Snow Cover, Fig. 2d), which likely improved habitat and breeding conditions.
In northwestern Alaska, large numbers of snow geese were observed along with cold spring conditions that would have been considered “normal” spring weather 30 years ago. Geese were present but food availability was limited by persistent snow and ice cover. The coast of the Yukon-Kuskokwim Delta (YKD; Fig. 1) was mostly snow free by mid-April, but lakes (see essay Lake Ice) and rivers were still frozen until mid-May. Migrating birds bypassed Bethel, Alaska, and arrived on the coast in early to mid-May. Warm and dry weather conditions during the nesting period (mid-May to June) may have negatively affected productivity. Decreased nesting effort (i.e., lower number of nests found) was observed on the YKD in 2022 for black brant, emperor geese, and cackling geese, with multiple research crews reporting loafing behavior and lack of nesting.
Local spring weather conditions and migratory bird observations are used each year by entities such as the Alaska Migratory Bird Co-Management Council (AMBCC; https://www.alaskamigratorybirds.com) to determine when to initiate spring harvest closures to protect birds and eggs during the primary nesting period (see Methods and data). Harvest dates were fixed prior to 2021, despite increasing variability in the phenology of spring weather and goose nesting behavior. The shift towards flexible start dates based on local observations is recent, representing widespread efforts to provide more effective evidence-based wildlife management through collaboration between management agencies and Indigenous community members in Alaska and Canada.
Population status of Arctic geese
Almost all North American populations of geese have been stable or have increased over the long-term, but about half of all Arctic and sub-Arctic populations have declined over the most recent 10-year period (Table 1). Population trends often determine management actions. For example, increased abundance of emperor geese allowed for an open harvest season in 2017 following a 30-year closure due to previously lower population numbers and Aleutian cackling geese were removed from the ‘endangered’ category of the Endangered Species List in 2001 in response to a dramatic increase in abundance (the 2022 population estimate was the highest on record). In contrast, Atlantic Population Canada geese, which breed primarily on the Ungava Peninsula (Fig. 1), have had restrictive harvest regulations in recent years due to declining abundance and low productivity, primarily due to multiple years of lower-than-average May temperatures. However, surveys in 2022 indicated improved breeding conditions and the first notable increase in the breeding population in almost a decade. For most northern-nesting goose populations, short-term declines have not led to management actions, because overall, most populations remain at or above historical levels.
| Species | Population | Survey Range | Growth Rate (% change/yr; All Survey Years) |
Growth Rate (% change/yr; Most Recent 10 Survey Years) |
|---|---|---|---|---|
| Brant | Atlantic | 1970-2022 | +1 | 0 |
| Pacific | 1970-2022 | 0 | 0 | |
| Cackling Goose | Midcontinent | 1987-2019 | +3 | -5 |
| Cackling | 1985-2022 | +4 | -6 | |
| Taverner’s | 2007-2022 | -1 | +4 | |
| Aleutian | 1975-2022 | +12 | +1 | |
| Canada | North Atlantic | 1990-2022 | 0 | -1 |
| Atlantic | 1993-2022 | +4 | -3 | |
| Southern Hudson Bay | 2016-2021 | 0 | 0 | |
| W. Prairie/Great Plains | 1970-2022 | +6 | +4 | |
| Dusky | 1986-2022 | 0 | -2 | |
| Lesser | 1970-2022 | -1 | +4 | |
| Emperor Goose | Yukon-Kuskokwim Delta | 1985-2022 | +2 | -2 |
| Snow Goose | Greater | 1970-2022 | +5 | -2 |
| Midcontinent Lesser | 1970-2019 | +6 | -6 | |
| Western Arctic Lesser | 2000-2018 | +3 | +11 | |
| Wrangel Island Lesser | 1975-2021 | +4 | +17 | |
| Ross’s | Western and Central Arctic | 1989-2019 | +9 | -2 |
| Greater white-fronted Goose | Pacific | 1985-2022 | +6 | -2 |
| Midcontinent | 1976-2020 | +4 | -3 |
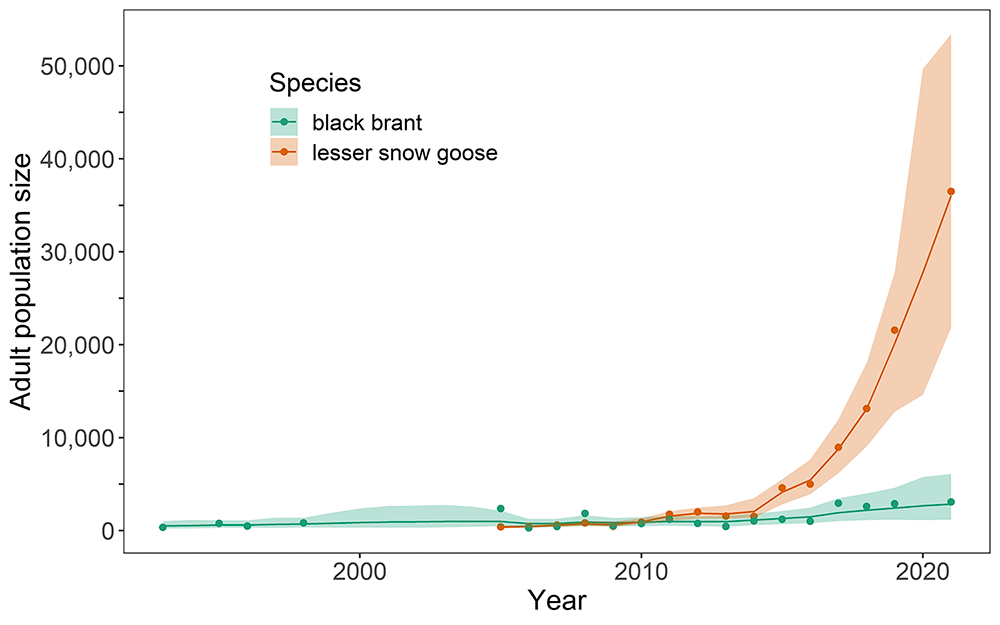
Abundance of Ross’s and greater and lesser snow geese in the midcontinent and eastern regions of North America has stabilized or declined in recent years. This recent change follows decades of exponential growth since the 1970s, which prompted overabundance designations and liberal harvest regulations beginning in 1999. Harvest probabilities (the proportion of the population likely harvested by hunters) have remained low with minimal impacts on population dynamics of Ross’s and midcontinent lesser snow geese (Alisauskas et al. 2022), but greater harvest probabilities were achieved on the smaller population of greater snow geese, which contributed to curtailing population growth (Lefebvre et al. 2017). Despite low harvest rates, biologists have documented declines in abundance at major snow goose breeding colonies in the midcontinent region, multiple years of poor productivity, high rates of emigration, and shifts in wintering and breeding distribution (Alisauskas et al. 2022; Weegman et al. 2022). These changes have been linked to resource limitations caused by climate change and negative feedbacks associated with high goose abundance. As a result, the midcontinent population of lesser snow geese has declined by approximately 60% since reaching peak abundance in 2007. In contrast, populations of lesser snow geese in the Western North American Arctic are exhibiting rapid growth due to improved climatic and habitat conditions in that region.
Observations from the Colville River Delta in Arctic Alaska distinctly highlight the effects of a rapidly warming Arctic on goose species. Both black brant and lesser snow geese have adjusted their behavior in tandem with phenological shifts, arriving earlier each spring and initiating nesting earlier in years with warmer springs and earlier snowmelt (Ruthrauff et al. 2021). Increasing goose populations on the Colville River Delta (Fig. 3), and elsewhere in Arctic Alaska, may also have benefited indirectly from permafrost thaw and coastal erosion, which have allowed nitrogen-rich salt-tolerant vegetation to replace less digestible freshwater meadows (Tape et al. 2013). Unlike black brant, however, snow geese can dig for belowground food when aboveground plant growth is delayed by cold weather. They also have a flexible breeding strategy, which allows them to be more successful breeders across years with variable spring conditions (Hupp et al. 2018). These traits may account for their explosive growth in the Western Arctic and on Wrangel Island (Table 1 and Fig. 3).
Climate and habitat changes in recent years have also altered distributions of geese during the non-breeding season. For example, large numbers of midcontinent greater-white-fronted, cackling, lesser snow, and Ross’s geese have moved eastward and northward from traditional wintering areas in coastal Texas and Louisiana, as agricultural production in those areas has decreased and winter temperatures have increased (VonBank et al. 2021). Black brant, which are a maritime goose, have substantially shifted their wintering distribution northward, with greater numbers overwintering at Izembek Lagoon and adjacent estuaries on the Alaska Peninsula, instead of at traditional wintering areas in Mexico (Olson 2021; Dave Safine, U.S. Fish and Wildlife Service, 2022, personal communication). Expansive drought in California and other western states is also altering goose distribution and migration patterns (Overton et al. 2021).
Methods and data
On the YKD and North Slope of Alaska, local observations are used to set a 30-day closure period of spring harvest, mandatory under the Migratory Bird Treaty Act (U.S. Fish and Wildlife Service 2020). As birds start arriving in spring, local observations are collected and communicated among hunters, tribal organizations, and observers, and the U.S. Fish and Wildlife Service (USFWS) contributes information about harvest and nest initiation that determine a range of likely closure dates. The final start date of the 30-day closure for the core nesting period is then communicated through radio, press release, and other methods.
Trends in Table 1 were calculated by regressing the natural logarithms of survey results on year. Estimates in Fig. 3 were derived using a Bayesian state-space model with lognormal process variation and log-normal observation errors. Points are observed counts, solid lines are the posterior mean estimates for each species, and shading represents the 95% posterior credible intervals.
Acknowledgments
Observations and perspectives of Arctic residents were sought and presented in this essay. Laura McDuffie (USGS), Dan Ruthrauff (USGS), Julian Fischer (USFWS), and two external reviewers provided helpful comments on earlier drafts. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
References
Alaska Division of Environmental Health, 2022: Highly pathogenic avian influenza (HPAI) outbreaks and biosecurity toolkit from USDA APHIS. Accessed 26 September 2022, https://dec.alaska.gov/eh/vet/announcements/avian-influenza-outbreaks/#resources.
Alisauskas, R. T., and Coauthors, 2022: Subpopulation contributions to a breeding metapopulation of migratory arctic herbivores: survival, fecundity and asymmetric dispersal. Ecography, 2022(7), e05653, https://doi.org/10.1111/ecog.05653.
Fox, A. D., and J. O. Leafloor (eds.), 2018: A global audit of the status and trends of Arctic and Northern Hemisphere goose populations. Conservation of Arctic Flora and Fauna International Secretariat: Akureyri, Iceland. ISBN 978-9935-431-66-0.
Hupp, J. W., D. H. Ward, D. X. Soto, and K. A. Hobson, 2018: Spring temperature, migration chronology, and nutrient allocation to eggs in three species of arctic-nesting geese: Implications for resilience to climate warming. Glob. Change Biol., 24, 5056-5071, https://doi.org/10.1111/gcb.14418.
Lefebvre, J., G., Gauthier, J. F., Giroux, A. Reed, E. T. Reed, and L. Bélanger, 2017: The greater snow goose Anser caerulescens atlanticus: Managing an overabundant population. Ambio, 46, 262-274, https://doi.org/10.1007/s13280-016-0887-1.
Olson, S. J., 2021: Pacific Flyway Data Book, 2021: U.S. Department of the Interior, Fish and Wildlife Service, Division of Migratory Bird Management, Vancouver, Washington.
Overton, C. T., and Coauthors, 2022: Megafires and thick smoke portend big problems for migratory birds. Ecology, 103(1), e03552, https://doi.org/10.1002/ecy.3552.
Parrett, J. P., T. Obritschkewitsch, and R. W. McNown, 2021: Avian studies for the Alpine Satellite Development Project, 2021, ABR, Inc. Available at: https://catalog.northslopescience.org/gl/dataset/2321.
Ruthrauff, D. R., V. P. Patil, J. W. Hupp, and D. H. Ward, 2021: Life-history attributes of Arctic-breeding birds drive uneven responses to environmental variability across different phases of the reproductive cycle. Ecol. Evol., 11, 18514-18530, https://doi.org/10.1002/ece3.8448.
Tape, K. D., P. L. Flint, B. W. Meixell, and B. V. Gaglioti, 2013: Inundation, sedimentation, and subsidence creates goose habitat along the Arctic coast of Alaska. Environ. Res. Lett., 8, 045031, https://doi.org/10.1088/1748-9326/8/4/045031.
U.S. Fish and Wildlife Service, 2020: Migratory bird subsistence harvest in Alaska; updates to the regulations: Federal Register, 85 FR 73233, FWS-R7-MB-2-2-0022.
U.S. Fish and Wildlife Service, 2022: Waterfowl population status, 2022. U.S. Department of the Interior, Washington, D.C. USA. https://www.fws.gov/library/collections/waterfowl-population-status-reports.
U.S. Geological Survey, 2022: Distribution of highly pathogenic avian influenza H5 and H5N1 in North America, 2021/2022. https://www.usgs.gov/media/images/distribution-highly-pathogenic-avian-influenza-h5-and-h5n1-north-america-20212022. Accessed 26 September 2022.
VonBank, J. A., M. D. Weegman, P. T. Link, S. A. Cunningham, K. J. Kraai, D. P. Collins, and B. M. Ballard, 2021: Winter fidelity, movements, and energy expenditure of midcontinent greater white-fronted geese. Mov. Ecol., 9, 2, https://doi.org/10.1186/s40462-020-00236-4.
Weegman, M. D., R. T. Alisauskas, D. K. Kellett, Q. Zhao, S. Wilson, and T. Telenský, 2022: Local population collapse of Ross’s and lesser snow geese driven by failing recruitment and diminished philopatry. Oikos, 2022(5), e09184, https://doi.org/10.1111/oik.09184.
November 22, 2022